IL-17在促进银屑病、银屑病关节炎和强直性脊柱炎(AS)等自身免疫疾病中起重要作用。截止到2024年8月27日,全球范围内共上市7款靶向IL-17/IL-17R的大分子生物药,包括6个IL-17单抗和1个IL-17RA单抗,分别是诺华的Secukinumab、礼来的Ixekizumab、Biocad的Netakimab、协和发酵麒麟的Brodalumab、优时比(UCB)的Bimekizumab、智翔金泰赛立奇单抗和恒瑞医药的夫那奇珠单抗。国内针对IL-17/IL-17R布局的药企还有君实、三生国健、康方、鑫康合/丽珠、荃信和华博等也在研发中。
表1. 靶向IL-17的抗体药物临床及临床前研发进展(来源于:药渡数据库)
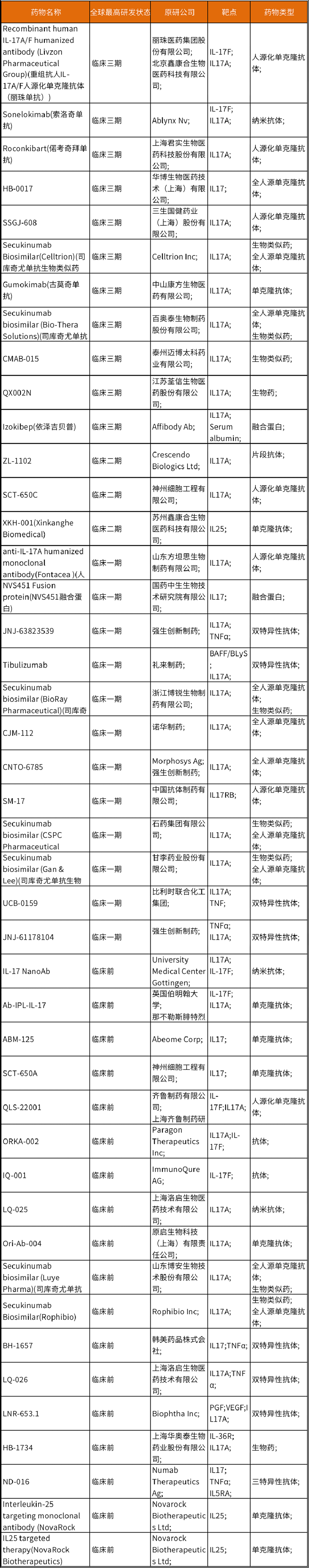
IL-17/IL-17R抗体药物研发如此之热,与IL-17的生物学功能密切相关。IL-17家族配体有六个亚型:IL-17A, IL-17B, IL-17C, IL-17 D, IL-17E(也叫做IL-25)和IL-17F。IL-17A和IL-17F形成同型二聚体和异型二聚体,而IL-17B、IL-17C和IL-17E形成同型二聚体[1]。IL-17R家族包括5个受体亚基,IL-17RA, IL-17RB, IL-17RC, IL-17RD和IL-17RE。IL-17RA是IL-17R家族最早发现的受体成员,是其他几种IL-17家族配体的共受体[2]。IL-17配体通过其对应的受体产生器官特异性的促炎或抗炎反应。
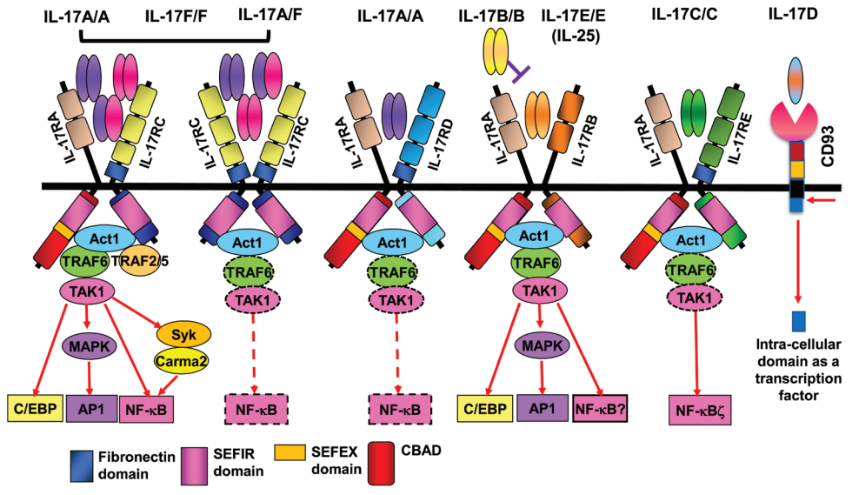
图1. IL-17因子和其受体家族[1]
IL-17配体家族
IL-17A
IL-17A在银屑病的发病机制中起核心作用,目前治疗此疾病的抑制性抗体主要也是针对此靶点。它作用于非造血细胞,特别是上皮细胞,并在邻近器官的免疫应答中发挥重要作用。在皮肤中,IL-17A导致角质细胞增殖和分化的改变,并诱导抗菌肽和趋化因子。IL-17A在其他炎症性疾病中也有作用,在多发性硬化症(MS)、类风湿关节炎和急性冠状动脉综合征患者中,IL-17A浓度升高与全身炎症参数相关[3]。
IL-17F
IL-17F和IL-17A被同一个基因位点(6p12) 编码,其调控方式相似。IL-17F可以与IL-17A形成异源二聚体。在银屑病患者中,IL-17F在斑块中的含量高于未受影响的皮肤部位。研究表明IL-17F和IL-17A具有协同作用:与单独抑制IL-17A不同,同时抑制这两种细胞因子可导致皮肤和关节成纤维细胞中炎症介质的下调显著增加。在细菌或真菌感染中IL-17F和IL-17A的功能类似[3]。
IL-17B
IL-17B在银屑病中的作用还不清楚,仅在银屑病皮损中有微弱表达。IL-17B不是由活化的T淋巴细胞产生的,但在中性粒细胞、B细胞、神经元细胞、基质细胞和结肠上皮细胞中都发现了IL-17B。类风湿性关节炎(RA)患者的滑膜和胰腺组织中IL-17B表达增加,IL-17B在成纤维细胞中增强TNF-α的作用,这对类风湿关节炎的免疫形成可能很重要[3]。
IL-17D
IL-17D在许多细胞和器官中表达,但在淋巴细胞和单核细胞等免疫细胞中表达微弱。IL-17D诱导内皮细胞产生IL-6、IL-8和GM-CSF。对骨髓前体细胞的造血也有抑制作用。其他研究表明IL-17D与病毒和肿瘤疾病有关,特别是通过自然杀伤细胞(NK细胞)的招募。然而,目前关于IL-17D在肿瘤发展过程中的数据是矛盾的。在动物模型中,IL-17D的缺乏导致了对病毒更高的易感性[3]。
IL-17C
IL-17C不是由T淋巴细胞产生,而是主要由上皮细胞如角质细胞产生。在人银屑病斑块中可以检测到高浓度的IL-17C。它可以作为上皮炎症的自分泌刺激物,特别是与TNF-α结合时。IL-17C在角质细胞中的过度表达导致小鼠银屑病样皮炎。IL-17C还能促进感觉神经的生长,在皮肤疱疹感染的小鼠模型中,IL-17C可以以类似于NGF(神经生长因子)的方式诱导神经元生长。也有报道称,在银屑病和特应性皮炎小鼠模型中,IL-17C的缺失显著降低了炎症反应。IL-17C因此作为免疫反应的上皮刺激物,独立于T细胞发挥作用[3]。
IL-17E (IL-25)
IL-17E诱导角质形成细胞增殖,并能够激活先天免疫细胞。在银屑病中,IL-17E由角质形成细胞强烈表达并激活特异性亚型巨噬细胞(M2或组织巨噬细胞)。银屑病皮损中IL-17E的表达与中性粒细胞数量呈正相关,与T淋巴细胞数量呈负相关。这是令人惊讶的,因为IL-17E在很长一段时间内被视为在Th2免疫应答中起作用的细胞因子。过敏性哮喘动物模型显示,用中和抗体抑制IL-17E可显著降低支气管高反应性和炎症反应中血清IgE浓度和组织学特征。同样,IL-17E在特应性湿疹中也有强烈表达[3]。
和IL-17C一样,IL-17E似乎能放大皮肤内固有的炎症过程,独立于适应性免疫系统的细胞[3]。
IL-17受体家族
图2. IL-17亚型与相关受体、产生细胞、对其他细胞的影响以及与疾病的关系概述[3]
IL-17信号通路
IL-17信号转导始于IL-17A/A、IL-17A/F或IL-17F/F细胞因子与其受体IL-17RA和IL-17RC的结合。与配体结合后,Act1激活多个通过不同TRAF蛋白介导的独立信号通路。TRAF6的激活可触发NF-κB、C/EBPβ、C/EBPδ和MAPK通路。IL-17R-Act1复合物也通过TRAF4与MEKK3和MEK5结合,诱发ERK5激活。TRAF6和TRAF4介导的IL-17信号通路可导致炎症基因的转录,而IL-17信号通路通过ACT1-TRAF2-TRAF5复合物控制IL-17靶基因mRNA的稳定性。因此,IL-17通过诱导新生基因转录或稳定靶基因mRNA转录来上调炎症基因表达[2]。
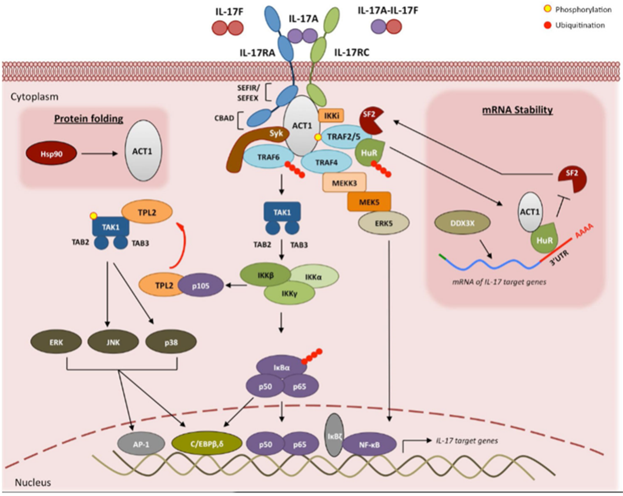
图4. IL-17信号通路[2]
近岸蛋白提供30余种IL-17系列靶点蛋白,适用于动物免疫、抗体筛选、功能评估和质量控制等不同环节的需求。
种类齐全:IL-17A/B/C/D/E/F、IL-17A&17F以及IL-17RA/RB/RD/RE共30余款靶点蛋白可根据需求任意选择;
不同标签:His/Fc/Biotin
高纯度:经过SDS-PAGE/SEC-HPLC验证
高活性:经过ELISA/BLI/Cell验证
部分数据展示:
经过SEC-HPLC验证:
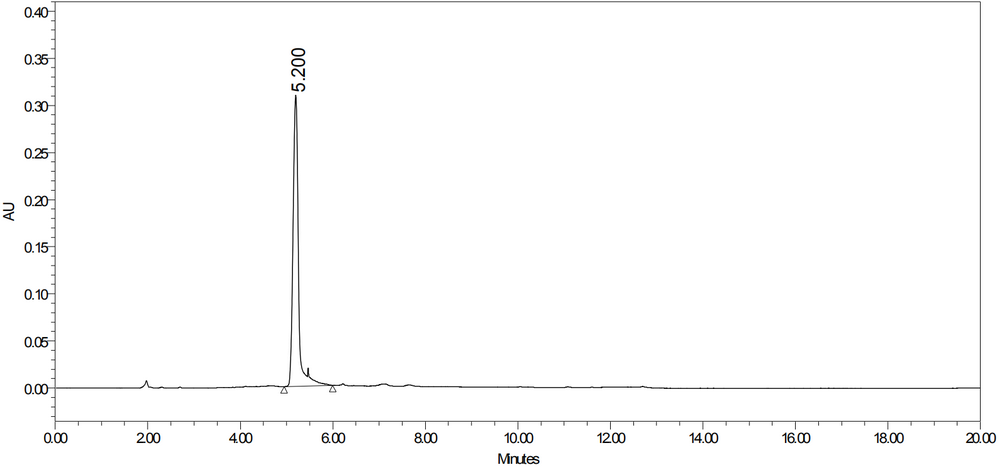 Greater than 95% as determined by SEC-HPLC. (Cat#C774)
Greater than 95% as determined by SEC-HPLC. (Cat#C774)
经过ELISA验证
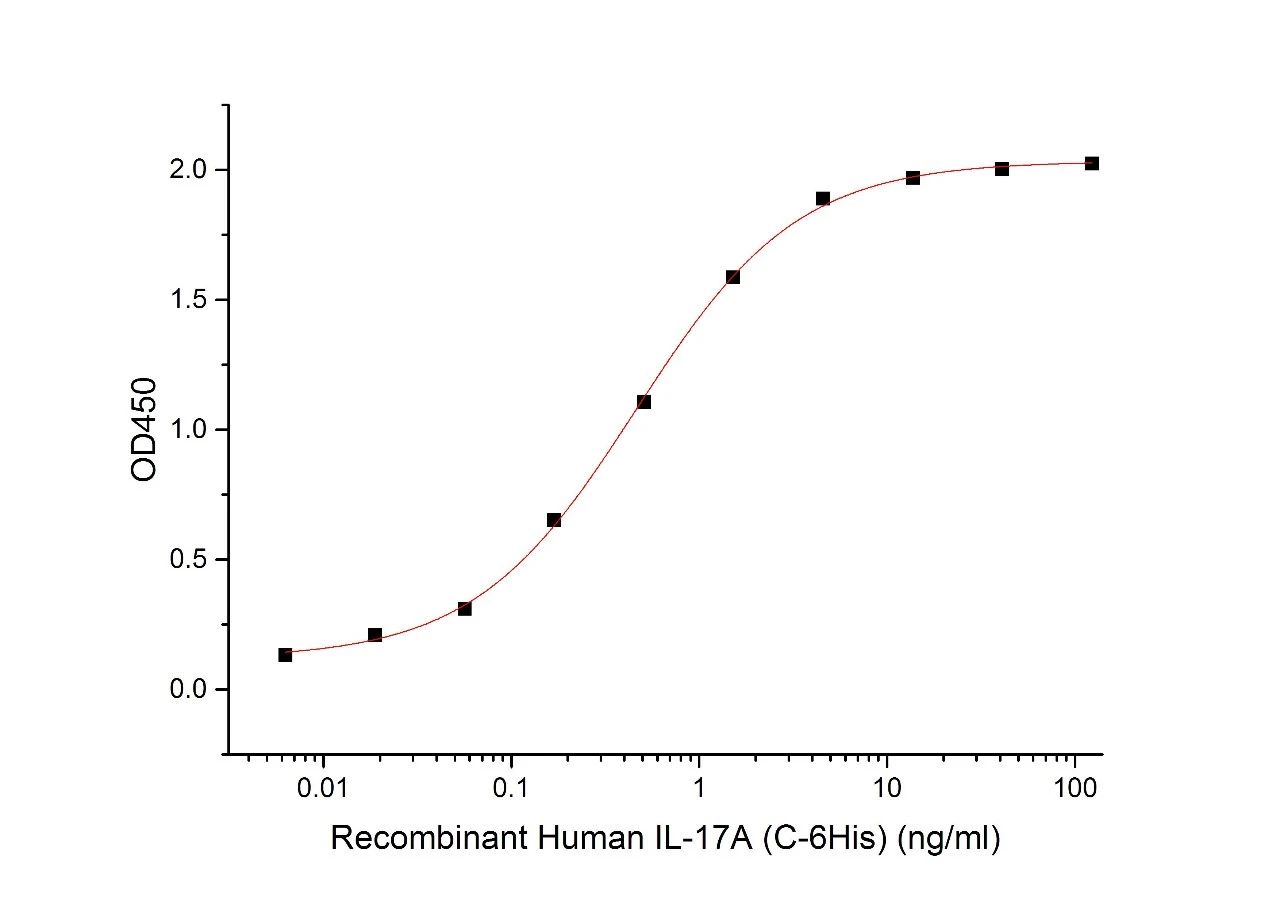
Immobilized Human IL-17RA-Fc(Cat#CI53) at 2μg/ml (100 μl/well)can bind Human IL-17A-His(Cat#C774).The ED50 of Human IL-17A-His(Cat#C774) is 0.49 ng/ml.
经过BLI验证
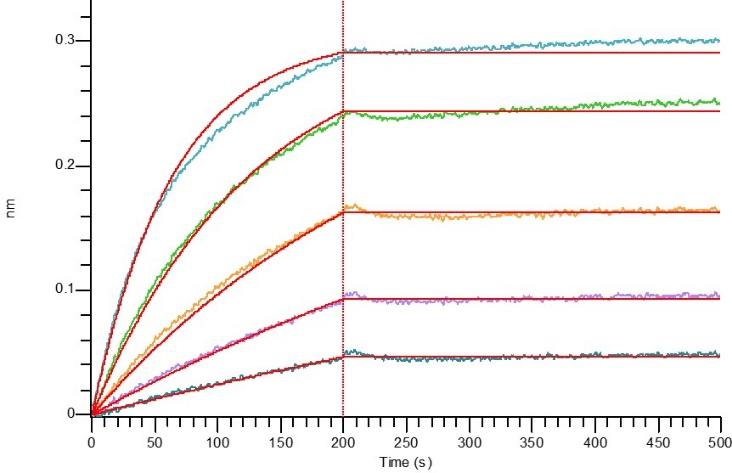
Loaded Biotinylated Human IL-17RA-Fc(Cat#CY89) on Pro A Biosensor, can bind Human IL-17A-His(Cat#C774) with an affinity constant of 0.003 nM as determined in BLI assay.
经过Cell验证
Measured by its ability to induce IL-6 secretion by NIH3T3 mouse embryonic fibroblast cells. The ED50 for this effect is 1-10 ng/ml.(Cat#C774)
相关产品:
|
Protein |
Product Name |
Cat. No. |
|
IL-17A |
Recombinant Human IL-17A,tag free |
C021 |
|
Recombinant Human IL-17A (C-6His) |
C774 |
|
|
Recombinant Mouse IL-17A (C-6His) |
CX14 |
|
|
Recombinant Cynomolgus IL-17A (C-6His) |
C02R |
|
|
IL-17B |
Recombinant Human IL-17B (C-Fc) |
CB78 |
|
IL-17C |
Recombinant Human IL-17C (C-6His) |
C13G |
|
Biotinylated Human IL-17C (C-6His-Avi) |
CY86 |
|
|
IL-17D |
Recombinant Human IL-17D,tag free |
CG96 |
|
IL-17E |
Recombinant Human IL-25 (C-6His) |
C792 |
|
IL-17F |
Recombinant Human IL-17F,tag free,E.coli |
C030 |
|
Recombinant Human IL-17F,tag free,Human Cells |
CS27 |
|
|
Recombinant Human IL-17F (N-6His) |
C14F |
|
|
Recombinant Human IL-17F (C-6His) |
CA22 |
|
|
Recombinant Mouse IL-17F (C-6His) |
CC11 |
|
|
Recombinant Cynomolgus IL-17F (C-6His) |
C12X |
|
|
IL-17A&17F |
Recombinant Human IL-17A&17F (C-6His) |
CI60 |
|
Biotinylated Human IL-17A&17F (C-Avi-6His) |
CY87 |
|
|
IL-17RA
|
Recombinant Human IL-17RA (C-6His) |
CU97 |
|
Recombinant Human IL-17RA (C-Fc) |
CI53 |
|
|
Biotinylated Human IL-17RA (C-Fc) |
CY89 |
|
|
Biotinylated Human IL-17RA (C-6His-Avi) |
CY78 |
|
|
Recombinant Mouse IL-17RA (C-Fc) |
C14H |
|
|
IL-17RB |
Recombinant Human IL-17RB (C-6His) |
C15B |
|
Recombinant Human IL-17RB (C-Fc) |
C12Y |
|
|
Recombinant Mouse IL-17RB (C-6His) |
C12U |
|
|
IL-17RD |
Recombinant Human IL-17RD (C-6His) |
C12K |
|
Recombinant Human IL-17RD (C-Fc) |
C12R |
|
|
Recombinant Mouse IL-17RD (C-6His) |
C12S |
|
|
Recombinant Mouse IL-17RD (C-Fc) |
C15U |
|
|
IL-17RE |
Recombinant Human IL-17RE (C-Fc) |
C13H |
|
Recombinant Mouse IL-17RE (C-6His) |
C13F |
|
|
Biotinylated Human IL-17RE (C-Fc-Avi) |
CY109 |
参考文献
[1] Chung, Soo-Hyun et al. “Interleukin-17 family members in health and disease.” International immunology vol. 33,12 (2021): 723-729. doi:10.1093/intimm/dxab075
[2] Amatya, Nilesh et al. “IL-17 Signaling: The Yin and the Yang.” Trends in immunology vol. 38,5 (2017): 310-322. doi:10.1016/j.it.2017.01.006
[3] Lauffer, Felix et al. “Cytokines of the IL-17 family in psoriasis.” Journal der Deutschen Dermatologischen Gesellschaft = Journal of the German Society of Dermatology : JDDG vol. 18,7 (2020): 675-681. doi:10.1111/ddg.14124